
You've reached the Virginia Cooperative Extension Newsletter Archive. These files cover more than ten years of newsletters posted on our old website (through April/May 2009), and are provided for historical purposes only. As such, they may contain out-of-date references and broken links.
To see our latest newsletters and current information, visit our website at http://www.ext.vt.edu/news/.
Newsletter Archive index: http://sites.ext.vt.edu/newsletter-archive/
A Newly Available Tool for Synchronization of Estrus in Swine
Livestock Update, December 2003
Mark Estienne, Swine Research Physiologist and Allen Harper, Extension Animal Scientist-Swine, Tidewater AREC
Introduction
Swine producers replace 30 to 50% of their sows with gilts each year (Gordon, 1997) and the introduction of females into the breeding herd is an important component of the overall reproductive efficiency of the operation. Large "gilt pools" are sometimes maintained to ensure that pubertal gilts will be in estrus and available for breeding at specified times. Reproductive efficiency would be enhanced if there were effective methods for synchronizing estrus in replacement gilts. Having replacement gilts exhibit estrus at a predicted time would allow more efficient scheduling and use of gestation and farrowing facilities and provide greater opportunity for introduction of females into sow groups when sows are bred following weaning.
Recently, the United States Food and Drug Administration (Federal Register, October 31, 2003) approved the use of altrenogest for synchronization of estrus in sexually mature gilts that have had at least one estrous cycle. Intervet Inc. (Millsboro, DE) currently markets altrenogest for use in horses under the trade name "Regu-mate". A 0.22% altrenogest solution for use in swine will now be marketed by Intervet under the trade name "Matrix". Matrix (6.8 ml containing 15 mg altrenogest/gilt) is administered once daily for 14 consecutive days by top-dressing on a portion of each gilt's daily feed. Gilts must not be slaughtered for human consumption for 21 days after the last treatment.
Herein we describe the physiology of the gilt estrous cycle and how and why altrenogest synchronizes estrus, and briefly review some research regarding the compound's efficacy.
Reproductive Physiology of the Gilt
Once gilts reach puberty at 6 to 8 months of age, they display estrus (synonyms = heat, sexual receptivity) at 18- to 22-day intervals throughout the year unless cycling is interrupted by pregnancy and lactation, poor nutrition, disease, etc. The physiological and behavioral changes associated with the estrous cycle are controlled by hormones.
The pituitary gland is a garden pea-sized organ that is located just below the brain in swine. Numerous hormones are secreted into the blood stream from the pituitary gland, including luteinizing hormone (LH) and follicle-stimulating hormone (FSH). Together LH and FSH are called gonadotropins.
Pituitary secretion of LH and FSH is ultimately controlled by the brain. Indeed, gonadotropin-releasing hormone (GnRH) is released from a specific area of the brain called the hypothalamus, travels through a microscopic system of blood vessels to the pituitary gland, and there stimulates LH and FSH secretion.
During the two to three day period just prior to estrus (i.e., proestrus), increasing blood levels of LH and FSH cause follicles to rapidly grow on each of the two ovaries. These follicles secrete increased levels of the hormone estrogen into the circulation, which in turn causes the behavioral and physiological changes associated with the two to three day-long estrus (e.g., reddening and swelling of the vulva, lordosis or the "standing response" in the presence of a boar, etc.). Moreover, each follicle contains an ovum (synonym = egg), which when fertilized by a sperm cell, develops into an embryo.
Normally, estrogen suppresses gonadotropin secretion. However, this ability weakens during proestrus. In fact, the rising concentrations of estrogen reach a threshold which triggers increased secretion of GnRH. This results in a massive release of LH occurring around the onset of estrus. The "LH surge" stimulates the process of ovulation. Multiple ova are released from the follicles on the ovaries during the process of ovulation, which occurs on average 40 hours after the onset of estrus. Ova are fertilized in the oviducts, which are tubes connecting the ovaries and uterus. The embryos then progress to the uterus and develop into fetuses.
The sites on the ovaries from which ova are ovulated subsequently form structures called corpora lutea that secrete another hormone, progesterone, into the circulation. During the luteal phase of the estrous cycle, progesterone, in concert with low levels of estrogen, inhibits LH and FSH secretion from the pituitary gland, and thus inhibits follicular growth. If fertilization and initiation of pregnancy did not occur during estrus, then the hormone prostaglandin-F2_ (PGF2_) is secreted into the circulation from the uterus around day 16 of the estrous cycle. PGF2_ causes the regression or death of the corpora lutea and as a result, progesterone levels decline. Decreasing levels of progesterone allow GnRH, and hence, LH and FSH, to increase, growth of follicles occurs, and the gilt continues to cycle.
Altrenogest: How and Why It Works
Altrenogest is an orally active progestin and has progesterone-like activity. When altrenogest is fed to a group of gilts, there is a suppression of gonadotropin secretion and as a consequence growth of follicles on the ovaries is abated. In terms of follicular development, ovaries remain in essentially a "quiescent" state for the duration of altrenogest treatment (usually 14 to 18 days). When altrenogest treatment is stopped, gonadotropin secretion increases and follicular growth ensues. Ultimately, gilts display synchronized estruses.
One may question why altrenogest must be administered for at least 14 days. This is because mature gilts will probably be randomly cycling and altrenogest feeding may commence when a portion of the gilts are in the luteal phase of the estrous cycle, with corpora lutea actively secreting progesterone. As mentioned above, the progesterone inhibits gonadotropin secretion and follicular development. These particular gilts must be given time for the corpora lutea to naturally regress. Otherwise, they would not show a synchronized estrus even though altrenogest feeding may have ceased.
Efficacy of Altrenogest for Synchronizing Estrus
In a number of research studies, altrenogest has been used to synchronize estrus in randomly cycling gilts (For reviews see Webel and Day, 1982; Day, 1984, Gordon, 1997). To summarize a number of studies, daily feeding of altrenogest (12.5 to 15 mg/day) for 14 to 18 days, synchronized estrus in approximately 90% of treated gilts. The onset of estrus after withdrawal of altrenogest was reported as 4 to 10 days with most gilts in estrus between 5 and 7 days.
In experiments conducted at the Tidewater Agricultural Research and Extension Center (TAREC) in Suffolk during the past few years, we have used altrenogest to successfully synchronize estrus in gilts (Estienne et al., 2001; Estienne and Harper, 2002; Horsley et al., 2004). For example, 86 randomly-cycling, crossbred gilts were given feed containing altrenogest (15 mg/day) for 18 days. As shown in Table 1, a large proportion of the gilts displayed estrus within 7 days after withdrawal of altrenogest. Figure 1 displays the degree to which estrus was synchronized following the end of altrenogest treatment.
After estrus, a large number of these gilts were slaughtered and the reproductive tracts examined. There appeared to be a normal ovulation rate, and the incidence of follicular and or luteal cysts was low (Table 1). Finally, a small group of altrenogest-treated gilts was mated via artificial insemination. At day 30 post-mating, 88.5% of the gilts (23/26) were confirmed pregnant and were carrying an average of 14.1 + 0.8 (mean + S.E.) live embryos.
Literature Cited
Estienne, M.J., A.F. Harper, B.R. Horsley, C.E. Estienne, and J.W. Knight. 2001. Effects of P.G. 600 on the onset of estrus and ovulation rate in gilts treated with Regu-mate. Journal of Animal Science 79:2757-2761.
Estienne, M.J. and A.F. Harper. 2002. Case study: Synchronization of estrus and fertility in gilts administered P.G. 600® after treatment with Regu-mate® for 14 or 18 days. The Professional Animal Scientist 18:158-161.
Day, B.N. 1984. Estrous cycle regulation. Proceedings of the 10th International Congress Animal Reproduction and AI (Urbana) IV, pp. 1-8.
Gordon, I. 1997. Controlled Reproduction in Pigs. CAB International, Wallingford, UK.
Horsley, B.R., M.J. Estienne, S.H. Purcell, H.K. Baitis, W.E. Beal, A.F. Harper, and J.W. Knight. Effect of P.G. 600 on the timing of ovulation in gilts treated with Regu-mate. Journal of Animal Science 82 (Supplement 1): (In Press).
Webel, S.K. and B.N. Day. 1982. The control of ovulation. In: D.J.A. Cole and G.R. Foxcroft (Eds.) Control of Pig Reproduction. Butterworths, London. pp. 197-210.
Table 1. Reproductive characteristics in gilts fed altrenogest (15 mg/d) for 18 days.
| Number of gilts | 86 |
| Gilts in estrus < 7 days after altrenogest withdrawal (%) | 93 (80/86) |
| Average days to estrus1 | 5.7 + 0.1 (range 4-7) |
| Gilts with follicular and/or luteal cysts (%) | 8.1 (6/74) |
| Average number of follicular and/or luteal cysts2 | 1.7 + 0.5 (range 1- 4) |
| Ovulation rate3 | 16.8 + 0.4 (range 11-26) |
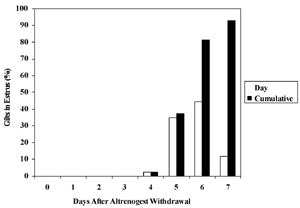
Figure 1. Daily and cumulative percentage of gilts (n = 86) in estrus after cessation of altrenogest treatment (15 mg/day for 18 days). Six gilts did not display estrus in < 7 days after withdrawal of altrenogest.