
You've reached the Virginia Cooperative Extension Newsletter Archive. These files cover more than ten years of newsletters posted on our old website (through April/May 2009), and are provided for historical purposes only. As such, they may contain out-of-date references and broken links.
To see our latest newsletters and current information, visit our website at http://www.ext.vt.edu/news/.
Newsletter Archive index: http://sites.ext.vt.edu/newsletter-archive/
Maximizing Boar Productivity with Optimum Trace Mineral Supplementation
Livestock Update, July 2005
Dr. Mark J. Estienne, Swine Research Physiologist and Dr. Allen F. Harper, Extension Animal Scientist-Swine, Tidewater AREC
This paper was presented on April 6, 2005 by Mark Estienne at the symposium entitled "Redefining Trace Mineral Nutrition: Supplementation strategies for modern diets and genetics", sponsored by Alltech in Nicholasville, KY.
Introduction
Trace minerals include iron, zinc, copper, selenium, molybdenum, manganese, chromium, cobalt, and iodine. Of these, zinc, copper, and selenium are generally considered as being important in male reproductive function (Bedwal and Bahuguna, 1994).
When compared to other species and even other classes of swine, research focusing on the trace mineral requirements of the breeding boar has been rather limited. Reasons for this relative lack of attention probably include the fact that boars did, and still do, comprise a relatively small part of the entire swine population. Additionally, long ago it was determined that a typical boar ejaculate contains many more sperm cells than are necessary to impregnate a single sow. Because natural mating systems dominated the industry, there was little incentive for investigating nutritional approaches for increasing the average number of fertile sperm cells produced in an ejaculate from say 50 billion to 75 billion. It was common for swine producers to feed boars a gestating sow diet and assume that male reproductive efficiency would not be seriously impacted. Today, however, artificial insemination is the most common mating system in the swine industry and each additional quality dose of semen processed from an ejaculate has monetary value.
Another factor that may have limited research in this area is the large variation displayed among boars with regard to reproductive characteristics such as semen volume, sperm concentration, sperm motility or measures of sexual behavior. To conduct meaningful research, detect statistically significant treatment differences, and draw sound conclusions, large numbers of experimental boars are generally needed which sometimes presents logistical problems for researchers. Moreover, when examining the effects of graded levels of trace minerals on reproduction in the boar, semen and libido characteristics may not be particularly sensitive measures. This may be in contrast to other swine nutritional research where a relatively small change in a specific nutrient results in easily demonstrated changes in blood chemistry, growth performance, or carcass quality.
The objective of this paper is to review some of the research that has been conducted to examine the effects of trace mineral supplementation on reproduction in the boar, with emphasis placed on zinc and selenium.
Control of Reproduction in Boars
Before delving into the topic of this paper, a brief review of boar reproductive physiology is in order. Shown in Figure 1 is the hormonal control of reproduction in boars. Gonadotropin-releasing hormone (GnRH) is released from the hypothalamus of the brain. GnRH travels to the pituitary gland and there stimulates the release of luteinizing hormone (LH) and follicle stimulating hormone (FSH), which travel via the circulation to the testes. In boars, LH and FSH stimulate the secretion of testosterone from the Leydig cells and estradiol from the Sertoli cells. These steroid hormones are together responsible for maintenance of libido. LH and FSH also stimulate the production of gametes (i.e., spermatogenesis). Spermatogenesis occurs in the Sertoli cells lining the seminiferous tubules of the testes, and is a process involving cell division and differentiation. Spermatogonia mitotically divide producing a set of spermatogonia and ultimately primary spermatocytes. These latter cells undergo meiotic divisions, first producing secondary spermatocytes that then become spermatids. The spermatids then differentiate into spermatozoa. In boars, spermatogenesis requires 6 to 7 weeks.
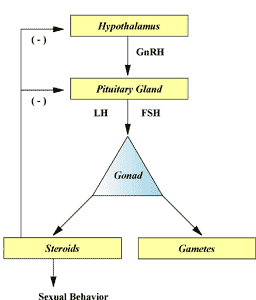
Figure 1. Hormonal control of reproduction in boars.
Evaluating Fertility in Boars
Both scientists and swine producers commonly evaluate semen in the laboratory, using assessments of the percentages of morphologically normal or motile spermatozoa as indicators of potential fertility of an ejaculate. Other, more sophisticated tests such as the hypoosmotic swelling test are used less frequently. With the hypoosmotic swelling test, spermatozoa with a biochemically active and functioning membrane will swell when hypoosmotically stressed due to the influx of water.
In reality, there currently are no laboratory tests that are completely reliable in assessing the potential fertility of an ejaculate. For example, at sperm motilities above 60%, no relationship existed between sperm motility and farrowing rates and number of pigs born live (Flowers, 1997). In other words, boars that consistently produce semen with 90% motile spermatozoa may be responsible for farrowing rates and litter sizes similar to those attributed to a boar that consistently produces semen with 60% motility. Nevertheless, the percentages of spermatozoa displaying progressive motility normal morphology, etc. can be used to identify ejaculates of overtly poor quality.
Trace Minerals in Sexually Active Boars
In 1998, the National Research Council (NRC) published the most recent Nutrient Requirements of Swine. Contained in Table 1 are the NRC requirements for some trace minerals in breeding boars. The table can be used to put into context, the results of experiments described below. With regard to selenium, it should be noted that current U.S. Food and Drug Administration (FDA) regulations allow up to 0.3 ppm of added selenium in the diets of all pigs (NRC, 1998).
Table 1. Trace mineral requirements of sexually active boars (from NRC, 1998).
| Feed intake | 2.0 kg/d Diet requirement (ppm) |
| Copper | 5 |
| Iodine | 0.14 |
| Iron | 80 |
| Manganese | 20 |
| Selenium | 0.15 |
| Zinc | 50 |
Zinc and Boar Productivity
Although zinc is considered essential for normal growth and reproduction, few studies have been conducted to investigate the effects of zinc supplementation on performance of boars. Boars fed diets deficient in zinc (22 to 29 ppm) displayed parakeratosis, and lower growth rates and feed intakes compared with boars fed diets with higher concentrations of zinc (48 to 53 ppm) (Liptrap et al., 1970).
Hesketh (1982) conducted an experiment during which Large White x Landrace boars were pair-fed diets containing zinc at levels of 22 to 25 ppm (n = 7) or 170 to 175 ppm (n = 7). The diets were fed for 7 to 8 weeks beginning when boars weighed 20 to 30 kg. Testicular weights were similar between groups but the zinc content of the testes was lower for the zinc deficient diet. There was no atrophy of the seminiferous tubules and germinal epithelium in the zinc-deficient boars but Leydig cells were smaller, less well-defined and contained an unusually high number of fat droplets. Lei et al. (1976) reported increased LH secretion, but a decreased testosterone response, after exogenous treatment with GnRH in zinc-deficient rats.
Liao et al. (1985) fed Duroc boars (n = 12) diets containing approximately 32, 89, 146 or 197 ppm zinc at a rate of 2 kg/head/d from 11 to 24 mo of age. Semen was collected every 4 to 5 d. There was no effect of treatment on the percentage of spermatozoa with morphological abnormalities but total spermatozoa produced was highest for boars fed diets supplemented with 89 or 146 ppm zinc. The authors suggested that the zinc concentration in the diet of working boars should be between 80 and 150 ppm, concentrations that are higher than current NRC recommendations.
Selenium and Boar Productivity
Selenium is essential for the normal development of spermatozoa and is incorporated into the mitochondrial capsula protein. Selenium is also a component of glutathionine peroxidase, an enzyme that protects cellular components against free radicals and is an antioxidant for cellular membrane lipids (Hansen and Deguchi, 1996). Generally, research has shown that selenium supplementation in boars increased glutathionine peroxidase activity and selenium concentrations in whole semen, spermatozoa, and seminal plasma, as well as in the circulation, kidney, liver, heart, skeletal muscle, testis, epididymis, prostate, seminal vesicle, and bulbourethral gland (Marin-Guzman et al., 1997; Segerson et al., 1981). However, Kolodziej and Jacyno (2004) reported that addition of selenium increased the concentration of selenium, but decreased glutathionine peroxidase activity, in seminal plasma. The amounts of selenium in seminal plasma of boars receiving either organic selenium or inorganic selenium were similar however glutathionine peroxidase activity favored the inorganic selenium source (Jacyno et al., 2002).
Effects of selenium supplementation on growth performance of boars. In general, selenium supplementation had little effect on the growth performance of boars. Indeed, growth rates, feed intakes, feed conversion efficiencies, and testicular sizes were similar for control boars and boars provided extra selenium in the diet (0.5 ppm; Kolodziej and Jacyno, 2004; Marin-Guzman, 1997) or via s.c. injections at 14-d intervals (Segerson et al., 1981).
In contrast to these reports, Henson et al. (1983) reported that boars fed diets supplemented with selenium exhibited some signs of retarded growth and sexual development. In that study, boars were fed a basal corn and soybean meal diet (0.05 ppm selenium) or the basal diet supplemented with a selenium premix at concentrations of either 0.10 (n = 11) or 0.25 ppm (n = 11). Experimental diets were fed on an ad libitum basis from 54 d of age until approximately 6 mo of age, and were then limit fed at a rate of 2.27 kg/boar/d. Treatment by age interactions existed for BW, testis width, libido (subjectively scored twice weekly upon exposure to ovariectomized, estrogen-treated gilts), and plasma testosterone levels, and in general values were greater for boars fed the basal diet than for those fed the basal diet supplemented with selenium.
Jacyno et al. (2002) conducted an experiment during which boars were fed diets supplemented with either 0.2 ppm organic selenium (selenium-yeast) and 60 ppm vitamin E (n = 40), or 0.2 ppm inorganic selenium (sodium selenite) and 30 ppm vitamin E (n = 40). Thus, source of selenium was confounded with concentration of vitamin E fed. The study was conducted from 70 to 180 d of age in both the summer and winter seasons, and the daily feed ration was gradually increased along with increasing BW. Compared with boars that received the organic source of selenium, boars receiving the inorganic selenium had higher average daily gain and better feed conversion efficiency, effects that were most pronounced during the winter. There were no effects of treatment on leanness or testicular size.
Effects of selenium supplementation on reproductive performance in breeding age boars. Several studies have been conducted to evaluate the effect of selenium supplementation on reproductive characteristics in breeding boars (Segerson et al. 1981; Marin-Guzman et al., 1997, 2000a, 2000b; Jacyno, 2002; Kolodziej and Jacyno, 2004).
Segerson et al. (1981) conducted an experiment during which crossbred boars were fed a low selenium diet (0.025 ppm) made of cornstarch and torula yeast on an ad libitum basis beginning at 78 d of age. Boars received s.c. injections of sodium selenite (0.33 mg/kg BW; n = 4) or 0.9% saline (n = 5) at 14-d intervals. At 230 d of age, boars were exposed to estrogen-treated, ovariectomized gilts and ejaculates were collected at 4- to 6-d intervals until a total of four ejaculates had been obtained from each boar. Injections of selenium increased the number of spermatozoa/ejaculate but had no effects on the percentages of viable or morphologically normal spermatozoa.
In a study involving 192 Landrace x Yorkshire x Duroc boars, Marin-Guzman et al. (1997) investigated the effects of dietary selenium and vitamin E supplementation on reproductive function. From weaning through 18 mo of age, boars were fed a basal diet made of torula yeast and dextrose or cornstarch (0.067 ppm selenium), the basal diet supplemented with sodium selenite (0.5 ppm), the basal diet supplemented with Vitamin E (220 IU/kg of diet), or the basal diet supplemented with both selenium (0.5 ppm) and Vitamin E (220 IU/kg of diet). Diets were consumed on an ad libitum basis from weaning to approximately 145 kg BW and thereafter were limit fed to individual boars at a rate of 2 kg/d.
In general, both dietary selenium and vitamin E enhanced boar semen quality, but the positive effects of added selenium on semen characteristics were more pronounced than were the effects of added vitamin E. During a 16-wk experimental period that began when boars were approximately 9 mo of age, semen was collected three times weekly. There were no effects of treatment on semen volume, sperm concentration or total spermatozoa. Sperm motility remained relatively constant when the diet was fortified with selenium but declined in boars fed the unfortified selenium diet. The percentage of normal spermatozoa declined in all treatment groups over the 16-wk period, but this decrease was least pronounced in boars receiving selenium supplementation. Boars fed the unfortified selenium diet had the highest percentages of spermatozoa with cytoplasmic droplets and bent and shoehook tails.
Subsequently, semen was collected twice weekly for 8 wk and at the end of this period, semen quality was again assessed. Supplementation with selenium increased semen volume, and the percentages of motile and morphologically normal spermatozoa. Gilts were artificially inseminated with semen from experimental boars and were killed 5 to 7 d later. Selenium supplementation increased fertilization rates and the number of accessory spermatozoa.
In other reports (Marin-Guzman et al., 2000a; 2000b), histological examination of testicular tissue of 18 mo-old boars employed in this study revealed that supplementation with selenium, but not vitamin E, increased the number of Sertoli cells, round spermatids, and secondary spermatocytes. Marin-Guzman et al. (2000a) suggested that selenium has a role in establishing the number of Sertoli cells and boar spermatozoal reserves. Also, selenium deficient boars produced spermatozoa with decreased ATP concentrations, and electron microscopy revealed that these cells had structural abnormalities to the tail midpiece, including altered mitochondrial shape and orientation and poor contact of the plasma membrane to the helical coil.
Jacyno et al. (2002) performed an experiment the objective of which was to compare organic versus inorganic selenium supplementation on semen characteristics in boars fed during either the winter or summer. In pigs, organic selenium is more effectively retained than inorganic selenium (Mahan and Parrett, 1996). Boars were fed diets supplemented with either 0.2 ppm organic selenium (selenium-yeast) and 60 ppm vitamin E (n = 40), or 0.2 ppm inorganic selenium (sodium selenite) and 30 ppm vitamin E (n = 40) beginning at 70 d of age. Beginning at 180 d of age, boars were trained to mount an artificial sow for semen collection. There was no effect of treatment on ejaculate volume or the percentage of motile spermatozoa. In contrast, the concentration of spermatozoa and total spermatozoa were higher in boars fed the diet containing the organic source of selenium. The quality of spermatozoa was also enhanced in boars receiving the organic selenium source. Indeed, boars fed the diet containing organic selenium had higher percentages of spermatozoa with normal acrosomes and that passed a hypoosmotic swelling test, and lower percentages of spermatozoa with minor or major morphological abnormalities. Beneficial effects of the organic selenium source were most evident in boars fed during the winter. Although these data are consistent with the notion that supplementation of diets with an organic source of selenium is superior to supplementation with inorganic selenium, source of selenium in this study was confounded with concentration of vitamin E.
Recently, Kolodziej and Jacyno (2004) evaluated semen characteristics in boars fed diets containing either 0.2 ppm selenium and 30 ppm vitamin E (n = 20), or 0.5 ppm selenium and 60 ppm vitamin E (n = 20), beginning at 70 d of age. Beginning at 180 d of age, boars were trained to mount an artificial sow and allow semen collection. There was no effect of treatment on gel-free semen volume or the percentage of motile spermatozoa. In contrast, the concentration of spermatozoa and total spermatozoa were higher in boars fed the diet containing the higher concentration of selenium and vitamin E. Boars fed the diet containing the higher concentration of selenium and vitamin E had higher percentages of spermatozoa with normal acrosomes and that passed a hypoosmotic swelling test, and lower percentages of spermatozoa with minor or major morphological abnormalities. While positive effects of a higher concentration of both dietary selenium and vitamin E were demonstrated, the relative contribution of each substance toward enhanced semen quality cannot be ascertained.
Few studies have evaluated the effects of trace mineral and vitamin supplementation on libido in boars. It is noteworthy that Kolodziej and Jacyno (2004) reported no effects of the higher concentration of selenium and vitamin E on the number of mounts of the artificial sow, the period from entering the collection area to the start of ejaculation, or the duration of ejaculation. In concert with these findings, Segerson et al. (1981) found no differences in the concentration of testosterone in serum of breeding boars of different selenium statuses. In contrast, boars fed a diet containing 0.2 ppm organic selenium and 60 ppm vitamin E mounted the artificial sow and began ejaculating more quickly, and had longer durations of ejaculations, than boars fed a diet containing 0.2 ppm inorganic selenium and 30 ppm Vitamin E (Jacyno et al., 2002).
Summary
Although research reports focusing on trace mineral supplementation of the sexually active boar are scarce, some general conclusions can be drawn. A prolonged period of restricted dietary zinc negatively impacts Leydig cell morphology and probably secretion of testosterone and limited research suggests that sperm produced per ejaculate is optimized at zinc concentrations between 80 and 150 ppm. With regard to reproductive performance, there is strong evidence to support the addition of selenium to boar diets. Improvements in sperm production, sperm morphology and actual fertility have been reported for boars fed diets supplemented with selenium generally at levels of 0.5 ppm. The FDA allows a maximum of only 0.3 ppm supplemental selenium in the diet of swine. Evidence suggests that organic sources of selenium may be more bio-available than inorganic selenium. Considering this, further research investigating the effects of organic source selenium on reproductive function in boars is warranted.
Literature Cited
Bedwal, R.S. and A. Bahuguna. 1994. Zinc, copper and selenium in reproduction. Experientia 50:626-640.
Flowers, W.L. 1997. Management of boars for efficient semen production. J. Reprod. Fertil. Suppl. 52:67-78.
Hansen, J.C. and Y. Deguchi. 1996. Selenium and fertility in animals and man- a review. Acta. Vet. Scand. 37:19-30.
Henson, M.C., H.G. Kattesh, J.P. Hitchcock, and S.A. Kincaid. 1983. The effects of dietary selenium on growth and selected reproductive parameters in young boars. Anim. Prod. 37:401-407.
Hesketh, J.E. 1982. Effects of dietary zinc deficiency on leydig cell ultrastructure in the boar. J. Comp. Path. 92:239-247.
Jacyno, E., M. Kawecka, M. Kamyczek, A. Kolodziej, J. Owsianny, and B. Delikator. 2002. Influence of inorganic SE + vitamin E and organic SE + vitamin E on reproductive performance of young boars. Agric. Food. Sci. Finland. 11:175-184.
Kolodziej, A. and E. Jacyno. 2005. Effect of selenium and vitamin E supplementation on reproductive performance of young boars. Archiv fur Tierzucht 48:68-75.
Lei, K.Y., A. Abbasi, and A.S. Prasad. 1976. Function of pituitary-gonadal axis in zinc deficient rats. J. Phys. 230:1730-1732.
Liao, C.W., S.C. Chyr, and T.F. Shen. 1985. The effect of dietary zinc content on reproductive performance of the boars. In: Proc. of the Third EAAP Animal Science Congress, Seoul, Korea Republic, 2:613-615.
Liptrap, D.O., E.R. Miller, D.E. Ullrey, D.L. Whitenack, B.L. Schoepke, and R.W. Luecke. 1970. Sex influence on the zinc requirement of developing swine. J. Anim. Sci. 30:736-741.
Mahan, D.C., and N.A. Parrett. 1996. Evaluating the efficacy of selenium-enriched yeast and sodium selenite on tissue selenium retention and serum glutathione peroxidase activity in grower and finisher swine. J. Anim. Sci. 74:2967-2974.
Marin-Guzman, J., D.C. Mahan, Y.K. Chung, J.L Pate, and W.F. Pope. 1997. Effects of dietary selenium and vitamin E on boar performance and tissue responses, semen quality, and subsequent fertilization rates in mature gilts. J. Anim. Sci. 75:2994-3003.
Marin-Guzman, J., D.C. Mahan, and J.L. Pate. 2000a. Effect of dietary selenium and vitamin E on spermatogenic development in boars. J. Anim. Sci. 78:1537-1543.
Marin-Guzman, J., D.C. Mahan, and R. Whitmoyer. 2000b. Effect of dietary selenium and vitamin E on the ultrastructure and ATP concentration of boar spermatozoa, and the efficacy of added sodium selenite in extended semen on sperm motility. J. Anim. Sci. 78:1544-1550.
NRC. 1998. Nutrient requirements of swine. 10th ed. Natl. Acad. Press, Washington, DC.
Segerson, E.C., W.R. Getz, and B.H. Johnson. 1981. Selenium and reproductive function in boars fed a low selenium diet. J. Anim. Sci. 53:1360-1367.