
You've reached the Virginia Cooperative Extension Newsletter Archive. These files cover more than ten years of newsletters posted on our old website (through April/May 2009), and are provided for historical purposes only. As such, they may contain out-of-date references and broken links.
To see our latest newsletters and current information, visit our website at http://www.ext.vt.edu/news/.
Newsletter Archive index: http://sites.ext.vt.edu/newsletter-archive/
Feeding Sows for a Rapid Return to Estrus after Weaning
Livestock Update, November 1999
Mark Estienne, Swine Research Physiologist, and Allen Harper, Extension Animal Scientist-Swine, Tidewater AREC
Introduction
Many factors influence the reproductive efficiency of a swine operation. One example is non-productive days (NPDs). NPDs accumulate for females that are not pregnant or lactating and include the interval between weaning and first service in sows. According to the Pigtales record keeping system, NPDs, farrowing rate and pigs born alive per litter were, in order, the three most important factors determining if a herd ranked at the top or bottom of a productivity group (Stein, 1992).
In modern breeding units, it is expected that sows return to estrus within seven days after weaning. For sows that do not exhibit estrus promptly after weaning, NPDs accumulate, thus compromising reproductive efficiency.
There are numerous factors that influence the rapidity with which sows return to estrus after weaning, and subsequent reproductive performance. This article focuses on the effect of nutrition on the weaning-to-estrus interval in sows. Before delving into that topic, a brief review of sow reproductive physiology is presented.
The brain ultimately controls pituitary secretion of LH and FSH. Indeed, gonadotropin-releasing hormone (GnRH) is released from a specific area of the brain called the hypothalamus, travels through a microscopic system of blood vessels to the pituitary gland, and there stimulates LH and FSH secretion.
Estrous Cycle. During the two to three day period just prior to estrus (i.e., proestrus), increasing blood levels of LH and FSH cause follicles to rapidly grow on each of the two ovaries. These follicles secrete increased levels of the hormone estradiol into the circulation, which in turn causes the behavioral and physiological changes associated with the two to three day-long estrus (e.g., reddening and swelling of the vulva, lordosis or the "standing response" in the presence of a boar, etc.). Moreover, each follicle contains an ovum (synonym = egg), which when fertilized by a sperm cell, develops into an embryo.
Normally, estradiol suppresses gonadotropin secretion. However, this ability weakens during proestrus, which allows LH and FSH secretion to increase. The rising concentrations of estradiol, produced from the growing follicles, reach a threshold that actually triggers increased secretion of GnRH. This results in a massive release of LH occurring around the onset of estrus. The "LH surge" stimulates the process of ovulation. Multiple ova are released from the follicles on the ovaries during the process of ovulation, which occurs on average 40 hours after the onset of estrus.
In mated sows, ova are fertilized by sperm cells in the oviducts, which are tubes connecting the ovaries and uterus. The embryos then progress to the uterus and develop into fetuses.
The sites on the ovaries from which ova are ovulated subsequently form structures called corpora lutea that secrete another hormone, progesterone, into the circulation. Progesterone, in concert with low levels of estradiol secreted from small ovarian follicles, inhibits LH and FSH secretion from the pituitary, and thus inhibits follicle growth. If fertilization and initiation of pregnancy does not occur, then the hormone prostaglandin-F2a2( (PGF2a) is secreted into the circulation from the uterus around Day 16 of the estrous cycle. PGF2a causes the death of the corpora lutea and as a result, progesterone levels decline. Decreasing levels of progesterone allow GnRH, and hence, LH and FSH, to increase, growth of follicles ensues, and the sow continues to "cycle" at 18 to 22 day intervals.
Gestation and Farrowing. As mentioned previously, the sites on the ovaries from which ova are ovulated subsequently form structures called corpora lutea, which secrete progesterone. If fertilization occurs and pregnancy is initiated, then PGF2a is not released into the circulation around Day 16 of the estrous cycle. Corpora lutea are rescued and secrete high levels of progesterone into the blood stream throughout gestation. Progesterone inhibits contractions of the uterus and is essential for maintenance of pregnancy.
PGF2a, however, is involved in the normal sequence of events that culminates in farrowing. Around Day 114 of gestation, large amounts of PGF2a are released from the uterus causing the corpora lutea to die. Consequently, progesterone levels in the blood decrease, uterine contractions commence and expulsion of fetuses occurs.
Lactation. As noted above, progesterone and estradiol inhibit gonadotropin secretion. Therefore, decreasing concentrations of progesterone, as well as lowered levels of estradiol produced from placental membranes, allow LH and FSH levels to increase, which in turn stimulates growth of ovarian follicles. Britt (1996) termed this stage of lactation the hypergonadotropic phase. If the litter is allowed to nurse, there is a suckling-induced suppression of LH and FSH secretion and within three days, concentrations of these two hormones become low. As a consequence, the follicles regress and there is no ovulation.
Between Days three and 14 post-farrowing, levels of LH and FSH are extremely low in the circulation of suckled sows and ovaries are devoid of large follicles. Suckling exerts a suppressive effect on GnRH secretion via feedback through nerves leading from the teats to the spinal cord and ultimately to the brain. During this period of lactation, called the transitional phase (Britt, 1996), the amount of LH being synthesized and stored in the pituitary is increasing as is the ability of the hypothalamus to secrete GnRH.
Between days 14 and 21 post-farrowing (normalization phase, Britt [1996]), release of LH and FSH into the circulation increases and this increase continues until weaning. The removal of the suckling stimulus at weaning allows a further increase in gonadotropin secretion. Follicles rapidly grow and there is the corresponding rise in circulating levels of estradiol. Sows return to estrus and the estradiol triggers the surge of LH, resulting in ovulation.
It is apparent that feed consumption during lactation has a profound effect on the rapidity with which sows display estrus after weaning (Table 1). Reduced daily feed intake during lactation increases the weaning-to-estrus interval.
| |||||||||||||||||||||||||||||||||||
Reese et al. (1982) conducted an experiment during which protein and other nutrients were kept constant, but energy consumption was varied among lactating sow groups to be 8, 12 or 16 mega-calories per sow per day. Sows consuming 8 mega-calories per day lost 57 lb of body weight and 8.4 mm of backfat during lactation and only 65% of these females displayed estrus within 8 days after weaning. Sows consuming 12 mega-calories per day lost 50% as much weight and backfat and over 91% exhibited estrus by Day 8 after weaning. Sows consuming 16 mega-calories per day lost even less weight and backfat and 96% were in estrus within 8 days after weaning.
King and Dunkin (1986) fed groups of lactating sows diets that were high energy-high protein, high energy-low protein, low energy-high protein or low energy-low protein. Sows fed the diet high in energy and protein (13 mega-calories and 650 g of protein per day) had a weaning-to-estrus interval of less than 7 days. In contrast, sows fed the other diets required 15 days or longer to return to estrus. Thus, failure of sows to consume enough energy and/or protein during lactation will result in an extended weaning-to-estrus interval.
The delay in the return to estrus after weaning in nutritionally deprived sows in all likelihood involves a suppression of gonadotropin secretion. Tokach et al. (1992) reported a positive relationship between energy intake and LH secretion in sows as early as 14 days after farrowing. Sows that returned to estrus in less than 9 days after weaning on Day 28 of lactation had higher circulating concentrations of LH at 14, 21 and 28 days postpartum than sows that required more than 15 days to return to estrus.
The number of eggs ovulated during the first estrus after weaning is lower and embryo survival is reduced for sows that have restricted feed intakes during lactation (Aherne et al., 1985). In fact, the amount of feed consumed during lactation could be the most important factor influencing subsequent reproduction in early-weaned sows. Koketsu et al. (1996) examined daily feed intake records on nearly 11,000 lactating sows on 16 commercial farms. In sows that consumed over 12.3 lb of feed per day during lactation, there was no effect of age of pigs at weaning on subsequent farrowing rates. However, when sows consumed less than 9.2 lb of feed per day there was an effect of weaning age on reproductive performance. Sows weaned at 11 to 13 days after farrowing had a lower farrowing rate than sows weaned at 14 to 22 days of lactation. Moreover, in sows consuming an intermediate amount of feed per day (9.2 lb to 12.3 lb per day), farrowing rate gradually increased as weaning age of pigs increased.
Feed intake during lactation is influenced by numerous factors including parity, environmental temperature and level of feeding during gestation.
Parity. Multiparous sows generally consume more feed during lactation than do primiparous sows (Britt, 1996). This could partially explain the fact that multiparous sows respond better to early weaning systems than do primiparous sows. In a retrospective study conducted using 178,519 litter records obtained from 13 commercial herds, Mabry et al. (1996) found that decreasing lactation length had a negative influence on the weaning-to-estrus interval, and this effect was most pronounced in primiparous sows compared to multiparous sows.
Environmental Temperature. Sows that farrow in the summer and early fall often have an extended weaning-to-estrus interval and the delay is more pronounced in primiparous compared to multiparous sows. Sows lactating during the summer eat less and loose more backfat. Barb et al. (1991) reported lower levels of LH in sows exposed to temperatures of 86(F compared to sows maintained at temperatures of 72ºF.
Feed Intake during Gestation. It is well established that the greater the feed intake during pregnancy, the lower the feed intake during lactation. Cole (1989) reported that when feed intake during pregnancy exceeds 4.4 lb per day, feed intake during the subsequent lactation is greatly reduced.
During a recent experiment, we evaluated performance during lactation and the weaning-to-estrus interval in sows that differed in body condition. In our study, the last rib backfat thickness was determined for 24 primiparous, Yorkshire x Landrace sows using a Lean-meater® ultrasound unit (Renco Corporation, Minneapolis, MN). Based on backfat thickness, sows were classified as FAT (over 25 mm; n = 7), MEDIUM (20 to 25 mm; n = 8) or THIN (less than 20 mm; n = 9) immediately after farrowing. Body weight, backfat thickness and body condition score (1 to 5; 1 = emaciated, 5 = obese) were different (P < .001) among groups (Table 2).
| ||||||||||||||||||||||||||||||||||||||||||||||||||
Total litter size, number born alive and average pig weight were similar among groups (P > .1) and overall were approximately 11.1, 10.2, and 3.5 lb, respectively. Pigs were weaned at approximately 22 days of age. The number of pigs weaned (9.3) and average pig weight at Day 17 of lactation (12.5 lb) did not differ (P > .1) among groups, suggesting that milk production was similar for FAT, MEDIUM and THIN sows.
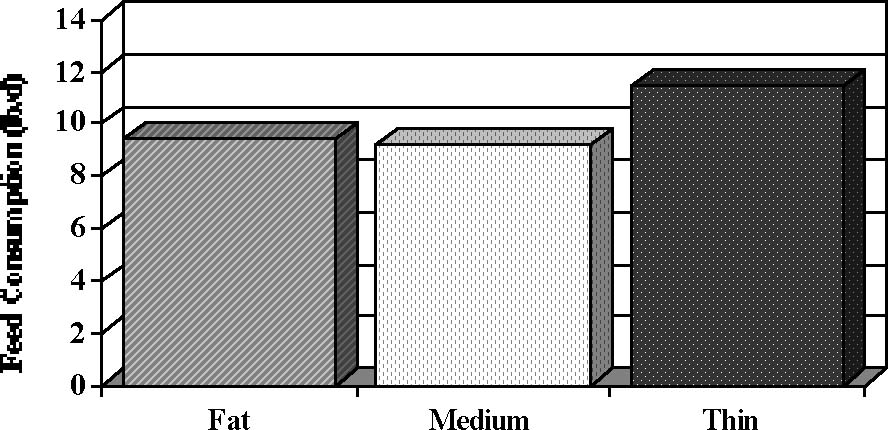
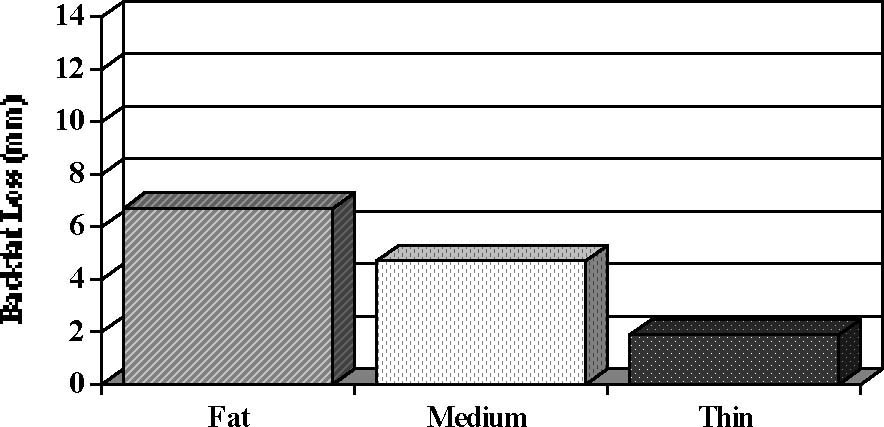
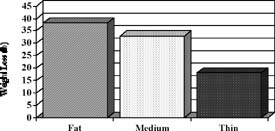
Weight loss (farrowing to Day 17 of lactation; P < .1), loss of backfat (farrowing to weaning; P < .004) and daily feed intake (farrowing to Day 17 of lactation; P < .09) differed among groups and are depicted in Figure 1.
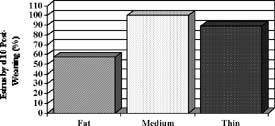
The percentage of sows displaying estrus within 10 days after weaning differed (P < .09) among groups (Figure 1). Among groups, sows displaying estrus during this period had a similar (P < .1) weaning-to-estrus interval (approximately 5.7 days). The data from our experiment suggest that over-conditioned sows had poorer rebreeding performance after weaning compared to thinner sows and this effect could be due to decreased feed consumption and greater weight and backfat loss during lactation.
 | 
|
 | 
|
Figure 1. Lactation performance and onset of estrus after weaning in sows of different body condition at farrowing (Estienne and Harper, unpublished data). At farrowing, sows were classified as FAT, MEDIUM or THIN based on last-rib backfat thickness. Pooled SE are .8 for feed consumption, .9 for backfat loss, and 6.3 for body weight loss.
In order to minimize the weaning-to-estrus interval, it is critical that a high plane of nutrition be achieved for the sow during lactation. The following are some key considerations for managers and herdsmen:
Aherne, F.X. and R.N. Kirkwood. 1985. Nutrition and sow prolificacy. J. Reprod. Fertil. Suppl. 33:169.
Barb, C.R., M.J. Estienne, R.R. Kraeling, D.N. Marple, G.B. Rampacek, C.H. Rahe, and J.L. Sartin. 1991. Endocrine changes in sows exposed to elevated ambient temperature during lactation. Domestic Anim. Endo. 8:117.
Britt, J.H. 1996. Biology and management of the early weaned sow. Proceedings of the American Association of Swine Practitioners 27th Annual Meeting pp. 417-426.
Cox, N.M., J.H. Britt, W.D. Armstrong, and H.D. Alhusen. 1983. Effect of feeding fat and altering weaning schedule on rebreeding in primiparous sows. J. Anim. Sci. 56:21.
Cole, D.J.A. 1989. Sow nutrition-The key to profitable pig production-more piglets in less time. Biotechnology in the Feed Industry- Proceedings of Alltech's Fifth Annual Symposium. pp. 107-120.
King, R.H. and A.C. Dunkin. 1986. The effect of nutrition on the reproductive performance of first-litter sows. 4. The relative effects of energy and protein intakes during lactation on the performance of sows and their piglets. Anim. Prod. 43:319.
Koketsu, Y., G.D. Dial, and V.L. King. 1996. Factors influencing farrowing rate on farms using early weaning. J. Anim. Sci. 74(Suppl. 1):31.
Mabry, J.W., M.S. Culbertson, and D. Reeves. 1996. Effects of lactation length on weaning to first service interval and first service farrowing rate in commercial sows. Swine Health and Production 4:185.
Pettigrew, J.E. and R.H. King. 1992. Modern sows need nutrition to match. National Hog Farmer 37(10):40.
Reese, D.E., B.D. Moser, E.R. Peo,Jr., A.J. Lewis, D.R. Zimmerman, J.E. Kinder, and W.W. Stroup. 1982. Influence of energy intake during lactation on the interval from weaning to first estrus in sows. J. Anim. Sci. 55:590.
Stein, T. 1992. Non-productive sow days: an overview. National Hog Farmer 37(10):6.
Tokach, M.D., J.E. Pettigrew, G.D. Dial, J.E. Wheaton, B.A. Crooker, and L.J. Johnston. 1992. Characterization of luteinizing hormone secretion in the primiparous, lactating sow: Relationship to blood metabolites and return-to-estrus interval. J. Anim. Sci. 70:2195.